Скелетные мышцы. Функции гладкой мышечной ткани. Гладкая мышечная ткань: строение Какими свойствами обладают гладкие мышцы
У позвоночных животных и человека различают три разных по строению группы мышц :
- поперечно-полосатые мышцы скелета;
- поперечно-полосатая мышца сердца;
- гладкие мышцы внутренних органов, сосудов и кожи.
Рис. 1. Виды мышц человека
Гладкие мышцы
Из двух видов мышечной ткани (поперечно-полосатой и гладкой) гладкая мышечная ткань находится на более низкой ступени развития и присуща низшим животным.
Образуют мышечный слой стенок желудка, кишечника, мочеточников, бронхов, кровеносных сосудов и других полых органов. Они состоят из веретенообразных мышечных волокон и не имеют поперечной исчерченности, так как миофибриллы в них расположены менее упорядоченно. В гладких мышцах отдельные клетки соединяются между собой специальными участками наружных мембран - нексусами . За счет этих контактов потенциалы действия распространяются с одного мышечного волокна на другое. Поэтому в реакцию возбуждения быстро вовлекается вся мышца.
Гладкие мышцы осуществляют движения внутренних органов, кровеносных и лимфатических сосудов. В стенках внутренних органов они, как правило, располагаются в виде двух слоев: внутреннего кольцевого и наружного продольного. В стенках артерии они формируют спиралевидные структуры.
Характерной особенностью гладких мышц является их способность к спонтанной автоматической деятельности (мышцы желудка, кишечника, желчного пузыря, мочеточников). Это свойство регулируется нервными окончаниями. Гладкие мышцы пластичны, т.е. способны сохранять приданную растяжением длину без изменения напряжения. Скелетная мышца, наоборот, обладает малой пластичностью и эту разницу легко установить в следующем опыте: если растянуть с помощью грузов и гладкую и поперечно-полосатую мышцы и снять груз, то скелетная мышца сразу же после этого укорачивается до первоначальной длины, а гладкая мышца долгое время может находиться в растянутом состоянии.
Такое свойство гладких мышц имеет большое значение для функционирования внутренних органов. Именно пластичность гладких мышц обеспечивает лишь небольшое изменение давления внутри мочевого пузыря при его наполнении.

Рис. 2. А. Волокно скелетной мышцы, клетка сердечной мышцы, гладкая мышечная клетка. Б. Саркомер скелетной мышцы. В. Строение гладкой мышцы. Г. Механограмма скелетной мышцы и мышцы сердца.
Гладким мышцам присущи те же основные свойства, что и поперечнополосатым скелетным мышцам, но и некоторые особые свойства:
- автоматия, т.е. способность сокращаться и расслабляться без внешних раздражений, а за счет возбуждений, возникающих в них самих;
- высокая чувствительность к химическим раздражителям;
- выраженная пластичность;
- сокращение в ответ на быстрое растяжение.
Сокращение и расслабление гладких мышц происходит медленно. Это способствует наступлению перестальтических и маятникообразных движений органов пищеварительного тракта, что приводит к перемещению пищевого комка. Длительное сокращение гладких мышц необходимо в сфинктерах полых органов и препятствует выходу содержимого: желчи в желчном пузыре, мочи в мочевом пузыре. Сокращение гладкомышечных волокон совершается независимо от нашего желания, под воздействием внутренних, не подчиненных сознанию причин.
Поперечно-полосатые мышцы
Поперечно-полосатые мышцы располагаются на костях скелета и сокращением приводят в движение отдельные суставы и все тело. образуют тело, или сому, поэтому их еще называют соматическими, а иннервирующую их систему — соматической нервной системой.
Благодаря деятельности скелетной мускулатуры осуществляется передвижение тела в пространстве, разнообразная работа конечностей, расширение грудной клетки при дыхании, движение головы и позвоночника, жевание, мимика лица. Насчитывается более 400 мышц. Общая масса мышц составляет 40% веса. Обычно средняя часть мышцы состоит из мышечной ткани и образует брюшко. Концы мышц — сухожилия построены из плотной соединительной ткани; они соединяются с костями при помощи надкостницы, но могут прикрепляться и к другой мышце, и к соединительному слою кожи. В мышце мышечные и сухожильные волокна объединяются в пучки при помощи рыхлой соединительной ткани. Между пучками располагаются нервы и кровеносные сосуды. пропорциональна количеству волокон, составляющих брюшко мышцы.

Рис. 3. Функции мышечной ткани
Некоторые мышцы проходят только через один сустав и при сокращении приводят его в движение — односуставные мышцы. Другие мышцы проходят через два или несколько суставов — многосуставные, они производят движение в нескольких суставах.
При концы мышцы, прикрепленные к костям, приближаются друг к другу, а размеры мышцы (длина) уменьшается. Кости, соединенные суставами, действуют как рычаги.
Изменяя положение костных рычагов, мышцы действуют на суставы. При этом каждая мышца влияет на сустав только в одном направлении. У одноосного сустава (цилиндрический, блоковидный) имеются две действующие на него мышцы или группы мышц, являющиеся антагонистами: одна мышца — сгибатель, другая — разгибатель. В то же время на каждый сустав в одном направлении действует, как правило, две мышцы и более, являющиеся синергистами (синергизм — совместное действие).
У двуосного сустава (эллипсоидный, мышелковый, седловидный) мышцы группируются соответственно двум его осям, вокруг которых совершаются движения. К шаровидному суставу, имеющему три оси движения (многоосный сустав), мышцы прилежат со всех сторон. Так, например, в плечевом суставе имеются мышцы-сгибатели и разгибатели (движения вокруг фронтальной оси), отводящие и приводящие (сагиттальная ось) и вращатели вокруг продольной оси, кнутри и кнаружи. Различают три вида работы мышц: преодолевающую, уступающую и удерживающую.
Если благодаря сокращению мышцы меняется положение части тела, то преодолевается сила сопротивления, т.е. выполняется преодолевающая работа. Работа, при которой сила мышцы уступает действию силы тяжести и удерживаемого груза, называется уступающей. В этом случае мышца функционирует, однако она не укорачивается, а удлиняется, например, когда невозможно поднять или удержать на весу тело, имеющее большую массу. При большом усилии мышц приходится опустить это тело на какую-нибудь поверхность.
Удерживающая работа выполняется благодаря сокращению мышц, тело или груз удерживается в определенном положении без перемещения в пространстве, например человек держит груз, не двигаясь. При этом мышцы сокращаются без изменения длины. Сила сокращения мышц уравновешивает массу тела и груза.
Когда мышца, сокращаясь, перемешает тело или его части в пространстве, они выполняют преодолевающую или уступающую работу, которая является динамической. Статистической является удерживающая работа, при которой не происходит движений всего тела или его части. Режим, при котором мышца может свободно укорачиваться, называется изотоническим (не происходит изменения напряжения мышцы и меняется только ее длина). Режим, при котором мышца не может укоротиться, называется изометрическим — меняется только напряжение мышечных волокон.

Рис. 4. Мышцы человека
Строение поперечно-полосатых мышц
Скелетные мышцы состоят из большого числа мышечных волокон, которые объединяются в мышечные пучки.
В одном пучке содержится 20-60 волокон. Мышечные волокна представляют собой клетки цилиндрической формы длиной 10-12 см и диаметром 10-100 мкм.
Каждое мышечное волокно имеет оболочку (сарколемму) и цитоплазму (саркоплазму). В саркоплазме находятся все компоненты животной клетки и вдоль оси мышечного волокна располагаются тонкие нити - миофибриллы, Каждая миофибрилла состоит из протофибрилл, в состав которых вкючены нити белков миозина и актина, являющихся сократительным аппаратом мышечного волокна. Миофибриллы разделены между собой перегородками, которые называются Z-мембранами, на участки - саркомеры. На обоих концах саркомеров к Z-мембране прикреплены тонкие актиновые нити, а в середине расположены толстые миозиновые нити. Нити актина своими концами частично входят между миозиновыми нитями. В световом микроскопе нити миозина выглядят в виде светлой полоски в темном диске. При электронной микроскопии скелетные мышцы выглядят исчерченными (поперечно-полосатыми).

Рис. 5. Поперечные мостики: Ак — актин; Мз — миозин; Гл — головка; Ш — шейка
На боковых сторонах миозиновой нити имеются выступы, получившие название поперечных мостиков (рис. 5), которые расположены под углом 120° по отношению к оси миозиновой нити. Актиновые филаменты выглядят в виде двойной нити, закрученной в двойную спираль. В продольных бороздках актиновой спирали находятся нити белка тропомиозина, к которым присоединен белок тропонин. В состоянии покоя молекулы белка тропомиозина расположены таким образом, чтобы предотвращать прикрепление поперечных мостиков миозина к актиновым нитям.

Рис. 6. А — организация цилиндрических волокон в скелетной мышце, прикрепленной к костям сухожилиями. Б — структурная организация филаментов в волокне скелетной мышцы, создающая картину поперечных полос.

Рис. 7. Строение актина и миозина
Во многих местах поверхностная мембрана углубляется в виде микротрубок внутрь волокна, перпендикулярно его продольной оси, образуя систему поперечных трубочек (Т-система). Параллельно миофибриллам и перпендикулярно поперечным трубочкам между миофибрилл расположена система продольных трубочек (саркоплазматический ретикулум). Концевые расширения этих трубочек - терминальные цистерны - подходят очень близко к поперечным трубочкам, образуя совместно с ними так называемые триады. В цистернах сосредоточено основное количество внутриклеточного кальция.
Механизм сокращения скелетной мышцы
Мышечная ткань состоит из клеток, называемых мышечными волокнами. Снаружи волокно окружено оболочкой — сарколеммой. Внутри сарколеммы содержится цитоплазма (саркоплазма), содержащая ядра и митохондрии. В ней содержится огромное количество сократительных элементов, называемых миофибриллами. Миофибриллы проходят от одного конца мышечного волокна до другого. Они существуют сравнительно короткий срок — около 30 суток, после чего и происходит их полная смена. В мышцах идет интенсивный синтез белка, необходимый для образования новых миофибрилл.
Мышечное волокно содержит большое количество ядер, которые располагаются непосредственно под сарколеммой, поскольку основная часть мышечного волокна занята миофибриллами. Именно наличие большого числа ядер обеспечивает синтез новых миофибрилл. Такая быстрая смена миофибрилл обеспечивает высокую надежность физиологических функций мышечной ткани.

Рис. 7. А — схема организации саркоплазматического ретикулума, поперечных трубочек и миофибрилл. Б — схема анатомической структуры поперечных трубочек и саркоплазматического ретикулума в индивидуальном волокне скелетной мышцы. В — роль саркоплазматического ретикулума в механизме сокращения скелетной мышцы
Каждая миофибрилла состоит из правильно чередующихся светлых и темных участков. Эти участки, обладая разными оптическими свойствами, создают поперечную исчерченность мышечной ткани.
В скелетной мышце сокращение вызывается поступлением к ней импульса по нерву. Передача нервного импульса с нерва на мышцу осуществляется через нервно-мышечный синапс (контакт).
Одиночный нервный импульс, или однократное раздражение, приводит к элементарному сократительному акту — одиночному сокращению. Начало сокращения не совпадает с моментом нанесения раздражения, поскольку существует скрытый, или латентный, период (интервал между нанесением раздражения и началом сокращения мышцы). В этот период происходит развитие потенциала действия, активация ферментных процессов и распад АТФ. После этого начинается сокращение. Распад АТФ в мышце приводит к превращению химической энергии в механическую. Энергетические процессы всегда сопровождаются выделением тепла и тепловая энергия обычно является промежуточной между химической и механическими энергиями. В мышце же химическая энергия превращается непосредственно в механическую. Но тепло в мышце образуется и за счет укорочения мышцы, и во время ее расслабления. Тепло, образующееся в мышцах, играет большую роль в поддержании температуры тела.
В отличие от сердечной мышцы, которая обладает свойством автоматики, т.е. она способна сокращаться под влиянием импульсов, возникающих в ней самой, и в отличие от гладкой мускулатуры, также способной к сокращению без поступления сигналов извне, скелетная мышца сокращается только при поступлении к ней сигналов из . Непосредственно сигналы к мышечным волокнам поступают по аксонам двигательных клеток, расположенным в передних рогах серого вещества спинного мозга (мотонейронам).
Рефлекторный характер деятельности мышц и координация мышечных сокращений
Скелетные мышцы в отличие от гладких способны совершать произвольные быстрые сокращения и производить этим значительную работу. Рабочим элементом мышцы является мышечное волокно. Типичное мышечное волокно представляет собой структуры с несколькими ядрами, отодвинутыми на периферию массой сократительных миофибрилл.
Мышечные волокна обладают тремя основными свойствами:
- возбудимостью — способностью отвечать на действия раздражителя генерацией потенциала действия;
- проводимостью — способностью проводить волну возбуждения вдоль всего волокна в обе стороны от точки раздражения;
- сократимостью — способностью сокращаться или изменять напряжение при возбуждении.
В физиологии имеется понятие двигательной единицы, под которой подразумевается один двигательный нейрон и все мышечные волокна, которые этот нейрон иннервирует. Двигательные единицы бывают разными по объему: от 10 мышечных волокон на единицу для мышц, выполняющих точные движения, до 1000 и более волокон на двигательную единицу для мышц «силовой направленности». Характер работы скелетных мышц может быть различным: статическая работа (поддержание позы, удержание груза) и динамическая работа (перемещение тела или груза в пространстве). Мышцы участвуют также в передвижении крови и лимфы в организме, выработке тепла, актах вдоха и выдоха, являются своеобразными депо для воды и солей, защищают внутренние органы, например мышцы брюшной стенки.
Для скелетной мышцы характерны два основных режима сокращения — изометрический и изотонический.
Изометрический режим проявляется в том, что в мышце во время ее активности нарастает напряжение (генерируется сила), но из-за того, что оба конца мышцы фиксированы (например, при попытке поднять очень большой груз), — она не укорачивается.
Изотонический режим проявляется в том, что мышца первоначально развивает напряжение (силу), способное поднять данный груз, а потом мышца укорачивается — меняет свою длину, сохраняя напряжение, равное весу удерживаемого груза. Чисто изометрического или изотонического сокращения практически наблюдать нельзя, но существуют приемы так называемой изометрической гимнастики, когда спортсмен напрягает мышцы без изменения длины. Эти упражнения в большей мере развивают силу мышц, чем упражнения с изотоническими элементами.
Сократительный аппарат скелетной мышцы представлен миофибриллами. Каждая миофибрилла диаметром 1 мкм состоит из нескольких тысяч протофибрилл — тонких, удлиненных полимеризированных молекул белков миозина и актина. Миозиновые нити в два раза тоньше актиновых, и в состоянии покоя мышечного волокна актиновые нити свободными кольцами входят между миозиновыми нитями.
В передаче возбуждения большую роль играют ионы кальция, которые входят в межфибриллярное пространство и запускают механизм сокращения: взаимное втягивание относительно друг друга актиновых и миозиновых нитей. Втягивание нитей происходит при обязательном участии АТФ. В активных центрах, расположенных на одном из концов миозиновых нитей, АТФ расщепляется. Энергия, выделяемая при расщеплении АТФ, преобразуется в движение. В скелетных мышцах запас АТФ невелик — всего на 10 одиночных сокращений. Поэтому необходим постоянный ре- синтез АТФ, который идет тремя путями: первый — за счет запасов креатинфосфата, которые ограничены; второй — гликолитический путь при анаэробном расщеплении глюкозы, когда на одну молекулу глюкозы образуется две молекулы АТФ, но одновременно образуется молочная кислота, которая тормозит активность гликолитических ферментов, и наконец третий — аэробное окисление глюкозы и жирных кислот в цикле Кребса, совершающееся в митохондриях и образующее 38 молекул АТФ на 1 молекулу глюкозы. Последний процесс наиболее экономичный, но очень медленный. Постоянная тренировка активизирует третий путь окисления, в результате чего повышается выносливость мышц к длительным нагрузкам.
Гладкие мышцы представлены в стенках органов пищеварительного канала, бронхов, кровеносных и лимфатических сосудов, мочевого пузыря, в матке, а также в радужной оболочке глаза, в цилиарной мышце, коже и железах. В отличие от поперечнополосатых мышц они не являются отдельными мышцами, а составляют только часть органов. Гладкие мышечные клетки имеют удлиненную веретенообразную или лентовидную форму с заостренными концами. Их длина у человека обычно бывает около 20 мкм. Наибольшей длины (до 500 мкм) достигают гладкие мышечные клетки в стенке беременной матки человека. В средней части клетки находится палочковидное ядро, а в цитоплазме вдоль всей клетки параллельно друг другу проходят тончайшие совершенно однородные миофибриллы. Поэтому клетка не имеет поперечной исчерченности. Более толстые миофибриллы расположены в наружных слоях клетки. Они называются пограничными и обладают одноосным двойным лучепреломлением. В электронном микроскопе видно, что миофибриллы являются пучками протофибрилл и обладают поперечной исчерченностью, не видимой в световом микроскопе. Гладкие мышечные клетки могут регенерировать путем деления (митоза). В них содержится разновидность актомиозина - тоноактомиозин. Между гладкими мышечными клетками имеются такие же участки контакта мембран, или нексусы, как и между сердечными, по которым, как предполагается, распространяется возбуждение и торможение с одних гладких мышечных клеток на другие.
В гладких мышцах возбуждение распространяется медленно Сокращения гладкой мышцы вызываются более сильными и более продолжительными раздражениями, чем скелетной. Латентный период ее сокращения продолжается несколько секунд. Гладкие мышцы сокращаются значительно медленнее скелетных. Так, период сокращения гладкой мышцы в желудке лягушки равен 15-20 с. Сокращения гладких мышц могут длиться многие минуты и даже часы. В отличие от скелетных мышц сокращения гладких мышц тонические. Гладкие мышцы способны при чрезвычайно малой затрате веществ и энергии долго находиться в состоянии тонического напряжения. Например, гладкие мышцы сфинктеров пищеварительного канала, мочевого пузыря, желчного пузыря, матки и других органов находятся в тонусе в течение десятков минут и многих часов. Гладкая мускулатура стенок кровеносных сосудов высших позвоночных животных остается в тонусе в течение всей жизни.
Существует прямая зависимость между частотой импульсов, возникающих в мышце, и уровнем ее напряжения. Чем больше частота, тем до известного предела больше тонус вследствие суммации напряжений неодновременно напрягающихся мышечных волокон.
Гладкие мышцы обладают тастичностью - способностью сохранять свою длину при растяжении без изменения напряжения в отличие от скелетных, которые при растяжении напряжены.
В отличие от скелетных мышц многие гладкие мышцы обладают автоматизмом. Они сокращаются под влиянием местных рефлекторных механизмов, например мейснеровского и ауэрбаховского сплетений в пищеварительном канале, или химических веществ, поступающих в кровь, например ацетилхолина, норадреналина и адреналина. Автоматические сокращения гладких мышц усиливаются или тормозятся под влиянием нервных импульсов, поступающих из нервной системы. Следовательно, в отличие от скелетных мышц существуют специальные тормозные нервы, которые прекращают сокращение и вызывают расслабление гладких мышц. Некоторые гладкие мышцы, имеющие большое количество нервных окончаний, не обладают автоматизмом, например сфинктер зрачка, мигательная перепонка кошки.
Гладкие мышцы могут сильно укорачиваться, значительно больше, чем скелетные. Одиночное раздражение может вызвать сокращение гладкой мышцы на 45%, а максимальное сокращение при частом ритме раздражения может достигать 60-75%.
Гладкая мышечная ткань развивается также из мезодермы (возникает из мезенхимы); она состоит из отдельных сильно вытянутых клеток веретенообразной формы, значительно меньшего размера по сравнению с волокнами поперечнополосатых мышц. Их длина колеблется от 20 до 500 μ, а ширина - от 4 до 7 μ. Как правило, эти клетки обладают одним лежащим в центре клетки удлиненной формы ядром. В протоплазме клетки в продольном направлении проходят многочисленные и очень тонкие миофибриллы, которые поперечной исчерченности не имеют и без особой обработки совершенно незаметны. Каждая гладкая мышечная клетка одета тончайшей соединительнотканной оболочкой. Этими оболочками соседние клетки связаны между собой. В отличие от поперечнополосатых волокон, расположенных почти во всю длину скелетной мышцы, на протяжении любого гладкомышечного комплекса встречается значительное число клеток, расположенных в одну линию.
Гладкие мышечные клетки встречаются в организме или разбросанными поодиночке в соединительной ткани, или связанными в мышечные комплексы различной величины.
В последнем случае каждая мышечная клетка бывает также окружена со всех сторон межклеточным веществом, пронизанным тончайшими фибриллами, количество которых может быть весьма различно. В межклеточном веществе обнаруживаются и тончайшие сети эластических волоконец.
Гладкие мышечные клетки органов объединяются в мышечные пучки. Во многих случаях (мочевые пути, матка и др.) эти пучки ветвятся и сливаются с другими пучками, образуя различной плотности поверхностные сети. Если же большое количество пучков располагается тесно, то образуется плотная мышечная оболочка (например, желудочно-кишечного тракта). Кровоснабжение гладких мышц осуществляется через сосуды, которые проходят в больших соединительнотканных прослойках между пучками; капилляры проникают между волокнами каждого пучка и, разветвляясь вдоль него, образуют густую капиллярную сеть. Гладкомышечная ткань содержит также лимфатические сосуды. Гладкие мышцы иннервируются волокнами вегетативной нервной системы. Гладкие мышечные клетки в отличие от волокон поперечнополосатых мышц производят медленные, длительные сокращения. Они способны работать долго и с большой силой. Например, мышечные стенки матки при родах, протекающих часами, развивают такую силу, которая недоступна для поперечнополосатых мышц. Деятельность гладких мышц, как правило, не подчинена нашей воле (вегетативная иннервация, см. ниже) - они непроизвольны.
Гладкая мускулатура по своему развитию (филогенезу) является более древней, чем поперечнополосатая, и в большей степени распространена у низших форм животного мира.
Классификация гладких мышц
Гладкие мышцы подразделяются на висцеральные (унитарные) и мультиунитарные. Висцеральные гладкие мышцы находятся во всех внутренних органах, протоках пищеварительных желез, кровеносных и лимфатических сосудах, коже. К мулыпиунитарным относятся ресничная мышца и мышца радужки глаза. Деление гладких мышц на висцеральные и мультиунитарные основано на различной плотности их двигательной иннервации. В висцеральных гладких мышцах двигательные нервные окончания имеются на небольшом количестве гладких мышечных клеток. Несмотря на это, возбуждение с нервных окончаний передается на все гладкие мышечные клетки пучка благодаря плотным контактам между соседними миоцитами - нексусам. Нексусы позволяют потенциалам действия и медленным волнам деполяризации распространяться с одной мышечной клетки на другую, поэтому висцеральные гладкие мышцы сокращаются одномоментно с приходом нервного импульса.
Функции и свойства гладких мышц
Пластичность . Еще одной важной специфической характеристикой гладкой мышцы является изменчивость напряжения без закономерной связи с ее длиной. Так, если растянуть висцеральную гладкую мышцу, то ее напряжение будет увеличиваться, однако если мышцу удерживать в состоянии удлинения, вызванным растяжением, то напряжение будет постепенно уменьшаться, иногда не только до уровня, существовавшего до растяжения, но и ниже этого уровня. Это свойство называется пластичностью гладкой мышцы. Таким образом, гладкая мышцы более похожа на тягучую пластичную массу, чем на малоподатливую структурированную ткань. Пластичность гладкой мускулатуры способствует нормальному функционированию внутренних полых органов.
Связь возбуждения с сокращением . Изучать соотношения между электрическими и механическими проявлениями в висцеральной гладкой мышце труднее, чем в скелетной или сердечной, так как висцеральная гладкая мышца находится в состоянии непрерывной активности. В условиях относительного покоя можно зарегистрировать одиночный ПД. В основе сокращения как скелетной, так и гладкой мышцы лежит скольжение актина по отношению к миозину, где ион Са2+ выполняет триггерную функцію.
В механизме сокращения гладкой мышцы имеется особенность, отличающая его от механизма сокращения скелетной мышцы. Эта особенность заключается в том, что прежде чем миозин гладкой мышцы сможет проявлять свою АТФазную активность, он должен быть фосфорилирован. Фосфорилирование и дефосфорилирование миозина наблюдается и в скелетной мышце, но в ней процесс фосфорилирования не является обязательным для активации АТФазной активности миозина. Механизм фосфорилирования миозина гладкой мышцы осуществляется следующим образом: ион Са2+ соединяется с кальмодулином (кальмодулин - рецептивный белок для иона Са2+). Возникающий комплекс активирует фермент - киназу легкой цепи миозина, который в свою очередь катализирует процесс фосфорилирования миозина. Затем происходит скольжение актина по отношению к миозину, составляющее основу сокращения. Отметим, что пусковым моментом для сокращения гладкой мышцы является присоединение иона Са2+ к кальмодулину, в то время как в скелетной и сердечной мышце пусковым моментом является присоединение Са2+ к тропонину.
Химическая чувствительность . Гладкие мышцы обладают высокой чувствительностью к различным физиологически активным веществам: адреналину, норадреналину, АХ, гистамину и др. Это обусловлено наличием специфических рецепторов мембраны гладкомышечных клеток. Если добавить адреналин или норадреналин к препарату гладкой мышцы кишечника, то увеличивается мембранный потенциал, уменьшается частота ПД и мышца расслабляется, т. е. наблюдается тот же эффект, что и при возбуждении симпатических нервов.
Норадреналин действует на α- и β-адренорецепторы мембраны гладкомышечных клеток. Взаимодействие норадреналина с β-рецепторами уменьшает тонус мышцы в результате активации аденилатциклазы и образования циклического АМФ и последующего увеличения связывания внутриклеточного Са2+. Воздействие норадреналина на α-рецепторы тормозит сокращение за счет увеличения выхода ионов Са2+ из мышечных клеток.
АХ оказывает на мембранный потенциал и сокращение гладкой мускулатуры кишечника действие, противоположное действию норадреналина. Добавление АХ к препарату гладкой мышцы кишечника уменьшает мембранный потенциал и увеличивает частоту спонтанных ПД. В результате увеличивается тонус и возрастает частота ритмических сокращений, т. е. наблюдается тот же эффект, что и при возбуждении парасимпатических нервов. АХ деполяризует мембрану, увеличивает ее проницаемость для Na+ и Са+.
Гладкие мышцы некоторых органов реагируют на различные гормоны. Так, гладкая мускулатура матки у животных в периоды между овуляцией и при удалении яичников относительно невозбудима. Во время течки или у животных, лишенных яичников, которым вводился эстроген, возбудимость гладкой мускулатуры возрастает. Прогестерон увеличивает мембранный потенциал еще больше, чем эстроген, но в этом случае электрическая и сократительная активность мускулатуры матки затормаживается.
Гладкие мышцы входят в состав внутренних органов. Благодаря сокращению они обеспечивают двигательную (моторную) функцию них органов (пищеварительный канал, мочеполовая система, кровеносные сосуды и т.д.). В отличие от скелетных мышц, гладкие мышцы являются непроизвольными.
Морфо-функциональная структура гладких мышц. Основной структурной единицей гладких мышц является мышечная клетка, которая имеет веретенообразную форму и покрыта снаружи плазматической мембраной. Под электронным микроскопом в мембране можно заметить многочисленные углубления - кавеолы, которые значительно увеличивают общую поверхность мышечной клетки. Сарколеммы непосмугованих мышечной клетки включает в себя плазматическую мембрану вместе с базальной мембраной, которая покрывает ее извне, и прилегающими коллагеновыми волокнами. Основные внутриклеточные элементы:ядро, митохондрии, лизосомы, микротрубочки, саркоплазматической сети и сократительные белки.
Мышечные клетки образуют мышечные пучки и мышечные слои. Межклеточное пространство (в 100 нм и более) заполнен эластичными и коллагеновыми волокнами, капиллярами, фибробластами и др.. В некоторых участках мембраны соседних клеток лежат очень плотно (щель между клетками составляет 2-3 нм). Предполагают, что эти участки (нексус) служат для межклеточного связи, передачи возбуждения. Доказано, что одни гладкие мышцы содержат большое количество нексус (сфинктер зрачка, циркулярные мышцы тонкой кишки и др.), у других их мало или совсем нет (семявыносящих протоков, продольные мышцы кишок). Между непосмугованих мышечными клетками существует также промежуточный, или десмоподибний, связь (через утолщение мембраны и с помощью отростков клеток). Очевидно, эти связи имеют значение для механического соединения клеток и передачи механической силы клетками.
Благодаря хаотичному распределению миозинових и актиновых протофибрилл клетки гладких мышц не поперечнополосатые, как скелетные и сердечная. В отличие от скелетных мышц, в гладких мышцах нет Т-системы, а саркоплазматической сети составляет только 2-7% объема миоплазмы и не имеет связей с внешней средой клетки.
Физиологические свойства гладких мышц .
Гладкомышечные клетки, - как-поперечнополосатые, сокращаются вследствие скольжения актиновых протофибрилл между миозиновои, однако скорость скольжения и гидролиз АТФ, а значит, и скорость сокращения, в 100-1000 раз меньше, чем в поперечнополосатых мышцах. Благодаря этому гладкие мышцы - хорошо приспособлены для длительного скольжения с небольшим затратой энергии и без усталости.
Гладкие мышцы с учетом способности генерировать ПД в ответ на пороговое или надгиорогове раздражение условно делят на фазные и тонические. Фазные мышцы генерируют полноценный ПД, тонические - только местный, хотя им присущ и механизм генерации полноценных потенциалов. Неспособность тонических мышц к ПД объясняется высокой калиевой проницаемостью мембраны, которая препятствует развитию регенеративной деполяризации.
Величина мембранного потенциала гладкомышечных клеток непосмугованих мышц варьирует от -50 до -60 мВ. Как и в других мышцах, в том числе и в нервных клетках, в его образовании принимают участие главным образом к +, Na +, Cl-. В гладкомышечных клетках пищеварительного канала, матки, некоторых сосудах мембранный потенциал нестабилен, наблюдаются спонтанные колебания в виде медленных волн деполяризации, на вершине которых могут появляться разряды ПД. Длительность ПД гладких мышц колеблется от 20-25 мс до 1 с и более (например, в мышцах мочевого пузыря), т.е. она длиннее, чем продолжительность ПД скелетных мышц. В механизме ПД гладких мышц рядом с Na + большую роль играет Са2 +.
Спонтанная миогенная активность. В отличие от скелетных мышц, гладкие мышцы желудка, кишок, матки, мочеточников имеют спонтанную миогенные активность, т.е. развивают спонтанные тетаногиодибни сокращения. Они хранятся в условиях изоляции этих мышц и при фармакологическом выключении интрафузальных нервных сплетений. Итак, ПД возникает в собственно гладких мышцах, а не обусловлен передачей в мышцы нервных импульсов.
Эта спонтанная активность имеет миогенные происхождения и возникает в мышечных клетках, которые выполняют функцию водителя ритма. В этих клетках местный потенциал достигает критического уровня и переходит в ПД. Но за реполяризацию мембраны спонтанно возникает новый местный потонциал, который вызывает еще один ПД, и т.д. ПД, распространяясь через нексус на соседние мышечные клетки со скоростью 0,05-0,1 м / с, охватывает весь мышцу, вызывая его сокращение. Например, перистальтические сокращения желудка возникают с частотой 3 раза за 1 мин, сегментарные и Маятникообразные движения толстой кишки-в 20 раз за 1 мин в верхних отделах и 5-10 за 1 мин - в нижних. Таким образом, гладкие мышечные волокна названных внутренних органов обладают автоматизмом, который проявляется их способностью ритмически сокращаться при отсутствии внешних раздражителей.
Какова причина возникновения потенциала в клетках гладких мышц водителя ритма? Очевидно, он возникает вследствие уменьшения калиевой и увеличение натриевой и кальциевой проницаемости мембраны. Что касается регулярного возникновения медленных волн деполяризации, наиболее выраженных в мышцах ЖКТ, го нет достоверных данных об их ионное происхождения. Возможно, определенную роль играет уменьшение первоначального инактивирующего компонента калиевого тока при деполяризации мышечных клеток вследствие инактивации соответствующих ионных калиевых каналов.
Эластичность и растяжимость гладких мышц. В отличие от скелетных мышц, гладкие при растяжении себя как пластичные, эластичные структуры. Благодаря пластичности гладкая мышца может быть полностью расслаблен как в сокращенном, так и в растянутыми состоянии. Например, пластичность гладких мышц стенки желудка или мочевого пузыря по мере наполнения этих органов предотвращает повышение внутриполостного давления. Чрезмерное растяжение часто приводит к стимулированию сокращения, которое обусловлено деполяризацией клеток водителя ритма, возникающий при растяжении мышцы, и сопровождается повышением частоты ПД, а вследствие этого - усилением сокращения. Сокращение, которое активизирует процесс растяжения, играет большую роль в саморегулировании базального тонуса кровеносных сосудов.
Механизм сокращения гладких мышц. Обязательным условием возникновения сокращение гладких мышц, как и скелетных, е увеличение концентрации Са2 + в миоплазми (до 10в-5 М). Считается, что процесс сокращения активизируется преимущественно внеклеточным Са2 +, поступающего в мышечные клетки через потенциалзависимые Са2 +-каналы.
Особенность нервно-мышечной передачи в гладких мышцах заключается в том, что иннервация осуществляется вегетативной нервной системой и она может оказывать как возбуждающий, так и тормозящее влияние. По типу различают холинергические (медиатор ацетилхолин) и адренергические (медиатор норадреналин) медиаторы. Первые обычно содержатся в мышцах пищеварительной системы, вторые - в мышцах кровеносных сосудов.
Один и тот же медиатор в одних синапсах может быть возбуждающих, а в других - тормозным (в зависимости от свойств циторецепторив). Адренорецепторы делят на а-и В-. Норадреналин, воздействуя на а-адренорецепторы, суживает кровеносные сосуды и тормозит моторику пищеварительного тракта, а воздействуя на В-адренорецепторы, стимулирует деятельность сердца и расширяет кровеносные сосуды некоторых органов, расслабляет мышцы бронхов. Описаны нервно-мышечно-. ную передачу в гладких мышцах за помощью и других медиаторов.
В ответ на действие возбуждающего медиатора происходит деполяризация клеток гладких мышц, которая проявляется в виде возбуждающего синаптической потенциала (ССП). Когда он достигает критического уровня, возникает ПД. Это происходит тогда, когда до нервного окончания друг за другом подходят несколько импульсов. Возникновение ЗСГИ является следствием увеличения проницаемости постсинаптической мембраны для Na +, Са2 + и СИ ".
Тормозной медиатор вызывает гиперполяризацию постсинаптической мембраны, что проявляется в тормозном синаптического потенциале (ГСП). В основе гиперполяризации лежит повышение проницаемости мембраны в основном для К +. Роль тормозного медиатора в гладких мышцах, возбуждаемые ацетилхолином (например, мышцы кишки, бронхов), играет норадреналин, а в гладких мышцах, для которых возбуждающих медиатором является норадреналин (например, мышцы мочевого пузыря), - ацетилхолин.
Клинико-физиологический аспект. При некоторых заболеваниях, когда нарушается иннервация скелетных мышц, их пассивное растяжение или смещение сопровождается рефлекторным повышением их тонуса, т.е. устойчивости к растяжению (спастичность или ригидность).
При нарушении кровообращения, а также под действием некоторых продуктов метаболизма (молочной и фосфорной кислот), ядовитых веществ, алкоголя, усталости, снижения температуры мышц (например, при длительном плавании в холодной воде) после длительного активного сокращения мышцы может возникать контрактура. Чем больше нарушается функция мышцы, тем сильнее выражена контрактурно последействие (например, контрактура жевательных мышц при патологии челюстно-лицевой области). Каково происхождение контрактуры? Считается, что контрактура возникла вследствие уменьшения в мышце концентрации АТФ, что привело к образованию постоянной связи между поперечными мостиками и актиновыми протофибрилл. При этом мышца теряет гибкость и становится твердым. Контрактура проходит, мышца расслабляется, когда концентрация АТФ достигает нормального уровня.
При заболеваниях типа миотонии клеточные мембраны мышц возбуждаются так легко, что даже незначительное раздражение (например, введение игольчатого электрода при электромиографии) обусловливает разряд мышечных импульсов. Спонтанные ПД (потенциалы фибрилляции) регистрируются также на первой стадии после денервации мышцы (пока бездействие не приведет к его атрофии).
Важным свойством гладкой мышцы является ее большая пластичность т. е. способность сохранять приданную растяжением длину без изменения напряжения. Различие между скелетной мышцей, обладающей малой пластичностью, и гладкой мышцей с хорошо выраженной пластичностью, легко обнаруживается, если их сначала медленно растянуть, а затем снять растягивающий груз. тотчас же укорачивается после снятия груза. В отличие от этого гладкая мышца после снятия груза остается растянутой до тех пор, пока под влиянием какого-либо раздражения не возникает ее активного сокращения.
Свойство пластичности имеет очень большое значение для нормальной деятельности гладких мышц стенок полых органов, например мочевого пузыря: благодаря пластичности гладкой мускулатуры стенок пузыря давление внутри него относительно мало изменяется при разной степени наполнения.
Возбудимость и возбуждение
Гладкие мышцы менее возбудимы, чем скелетные: их пороги раздражения выше, а хронаксия длиннее. Потенциалы действия большинства гладкомышечных волокон имеют малую амплитуду (порядка 60 мв вместо 120 же в скелетных мышечных волокнах) и большую продолжительность - до 1-3 секунд. На рис. 151 показан потенциал действия одиночного волокна мышцы матки.
Рефрактерный период продолжается в течение всего периода потенциала действия, т. е. 1-3 секунд. Скорость проведения возбуждения варьирует в разных волокнах от нескольких миллиметров до нескольких сантиметров в секунду.
Существует большое число различных типов гладких мышц в теле животных и человека. Большинство полых органов тела выстлано гладкими мышцами, имеющими сенцитиальный тип строения. Отдельные волокна таких мышц очень тесно примыкают друг к другу и создается впечатление, что морфологически они составляют единое целое.
Однакоэлектронномикроскопические исследования показали, что мембранной и протоплазматической непрерывности между отдельными волокнами мышечного синцития не существует: они отделены друг от друга тонкими (200-500 Å) щелями. Понятие «синцитиальное строение» является в настоящее время скорее физиологическим, чем морфологическим.
Синцитий - это функциональное образование, которое обеспечивает то, что потенциалы действия и медленные волны деполяризации могут беспрепятственно распространяться с одного волокна на другое. Нервные окончания расположены только на небольшом числе волокон синцития. Однако вследствие беспрепятственного распространения возбуждения с одного волокна на другое вовлечение в реакцию всей мышцы может происходить, если нервный импульс поступает к небольшому числу мышечных волокон.
Сокращение гладкой мышцы
При большой силе одиночного раздражения может возникать сокращение гладкой мышцы. Скрытый период одиночного сокращения этой мышцы значительно больше, чем скелетной мышцы, достигая, например, в кишечной мускулатуре кролика 0,25- 1 секунды. Продолжительность самого сокращения тоже велика (рис. 152 ): в желудке кролика она достигает 5 секунд, а в желудке лягушки - 1 минуты и более. Особенно медленно протекает расслабление после сокращения. Волна сокращения распространяется по гладкой мускулатуре тоже очень медленно, она проходит всего около 3 см в секунду. Но эта медленность сократительной деятельности гладких мышц сочетается с большой их силой. Так, мускулатура желудка птиц способна поднимать 1 кг на 1см2 своего поперечного сечения.
Тонус гладкой мышцы
Вследствие медленности сокращения гладкая мышца даже при редких ритмических раздражениях (для желудка лягушки достаточно 10-12 раздражений в минуту) легко переходит в длительное состояние стойкого сокращения, напоминающее тетанус скелетных мышц. Однако энергетические расходы при таком стойком сокращении гладкой мышцы очень малы, что отличает это сокращение от тетануса поперечнополосатой мышцы.
Причины, вследствие которых гладкие мышцы сокращаются и расслабляются много медленнее, чем скелетные, полностью еще не выяснены. Известно, что миофибриллы гладкой мышцы так же, как и скелетной мышцы, состоят из миозина и актина. Однако в гладких мышцах нет поперечной исчерченности, нет мембраны Z и они гораздо богаче саркоплазмой. По-видимому, эти особенности структуры гладких мышечных волн и обусловливают медленный темп сократительного процесса. Этому соответствует и относительно низкий уровень обмена веществ гладких мышц.
Автоматия гладких мышц
Характерной особенностью гладких мышц, отличающей их от скелетных, является способность к спонтанной автоматической деятельности. Спонтанные сокращения можно наблюдать при исследовании гладких мышц желудка, кишок, желчного пузыря, мочеточников и ряда других гладкомышечных органов.
Автоматия гладких мышц имеет миогенное происхождение. Она присуща самим мышечным волокнам и регулируется нервными элементами, которые находятся в стенках гладкомышечных органов. Миогенная природа автоматии доказана опытами на полосках мышц кишечной стенки, освобожденных путем тщательной препаровки от прилежащих к ней нервных сплетений. Такие полоски, помещенные в теплый растввр Рингера-Локка, который насыщается кислородом, способны совершать автоматические сокращения. При последующей гистологической проверке было обнаружено отсутствие в этих мышечных полосках нервных клеток.
В гладких мышечных волокнах различают следующие спонтанные колебания мембранного потенциала: 1) медленные волны деполяризации с длительностью цикла порядка нескольких минут и амплитудой около 20 мв; 2) малые быстрые колебания потенциала, предшествующие возникновению потенциалов действия; 3) потенциалы действия.
На все внешние воздействия гладкая мышца реагирует изменении частоты спонтанной ритмики, следствием которой являются сокращения и расслабления мышцы. Эффект раздражения гладкой мускулатуры кишки зависит от соотношения между частотой стимуляции и собственной частотой спонтанной ритмики: при низком тонусе - при редких спонтанных потенциалах действия - приложенное раздражение усиливает тонус при высоком же тонусе в ответ на раздражение возникает расслабление, так как чрезмерное учащение импульсации приводит к тому, что каждый следующий импульс попадает в рефрактерную фазу от предыдущего.
Гладкие мышцы входят в состав внутренних органов. Благодаря сокращению они обеспечивают двигательную (моторную) функцию них органов (пищеварительный канал, мочеполовая система , кровеносные сосуды и т.д.). В отличие от скелетных мышц, гладкие мышцы являются непроизвольными.
Морфо-функциональная структура гладких (не исполосованных) мышц.
Основной структурной единицей гладких мышц является мышечная клетка, которая имеет веретенообразную форму и покрыта снаружи плазматической мембраной. Под электронным микроскопом в мембране можно заметить многочисленные углубления - кавеолы, которые значительно увеличивают общую поверхность мышечной клетки. Сарколеммы непосмугованих мышечной клетки включает в себя плазматическую мембрану вместе с базальной мембраной, которая покрывает ее извне, и прилегающими коллагеновыми волокнами. Основные внутриклеточные элементы:
ядро, митохондрии, лизосомы, микротрубочки, саркоплазматической сети и сократительные белки.
Мышечные клетки образуют мышечные пучки и мышечные слои. Межклеточное пространство (в 100 нм и более) заполнен эластичными и коллагеновыми волокнами, капиллярами, фибробластами и др.. В некоторых участках мембраны соседних клеток лежат очень плотно (щель между клетками составляет 2-3 нм). Предполагают, что эти участки (нексус) служат для межклеточного связи, передачи возбуждения. Доказано, что одни гладкие мышцы содержат большое количество нексус (сфинктер зрачка, циркулярные мышцы тонкой кишки и др.), у других их мало или совсем нет (семявыносящих протоков, продольные мышцы кишок). Между непосмугованих мышечными клетками существует также промежуточный, или десмоподибний, связь (через утолщение мембраны и с помощью отростков клеток). Очевидно, эти связи имеют значение для механического соединения клеток и передачи механической силы клетками.
Благодаря хаотичному распределению миозинових и актиновых протофибрилл клетки гладких мышц не поперечнополосатые, как скелетные и сердечная. В отличие от скелетных мышц, в гладких мышцах нет Т-системы, а саркоплазматической сети составляет только 2-7% объема миоплазмы и не имеет связей с внешней средой клетки.
Физиологические свойства гладких мышц.
Гладкомышечные клетки, - как-поперечнополосатые, сокращаются вследствие скольжения актиновых протофибрилл между миозиновои, однако скорость скольжения и гидролиз АТФ, а значит, и скорость сокращения, в 100-1000 раз меньше, чем в поперечнополосатых мышцах. Благодаря этому гладкие мышцы - хорошо приспособлены для длительного скольжения с небольшим затратой энергии и без усталости.
Гладкие мышцы с учетом способности генерировать ПД в ответ на пороговое или надгиорогове раздражение условно делят на фазные и тонические. Фазные мышцы генерируют полноценный ПД, тонические - только местный, хотя им присущ и механизм генерации полноценных потенциалов. Неспособность тонических мышц к ПД объясняется высокой калиевой проницаемостью мембраны, которая препятствует развитию регенеративной деполяризации.
Величина мембранного потенциала гладкомышечных клеток непосмугованих мышц варьирует от -50 до -60 мВ. Как и в других мышцах, в том числе и в нервных клетках, в его образовании принимают участие главным образом к +, Na +, Cl-. В гладкомышечных клетках пищеварительного канала, матки , некоторых сосудах мембранный потенциал нестабилен, наблюдаются спонтанные колебания в виде медленных волн деполяризации, на вершине которых могут появляться разряды ПД. Длительность ПД гладких мышц колеблется от 20-25 мс до 1 с и более (например, в мышцах мочевого пузыря), т.е. она
длиннее, чем продолжительность ПД скелетных мышц. В механизме ПД гладких мышц рядом с Na + большую роль играет Са2 +.
Спонтанная миогенная активность.
В отличие от скелетных мышц, гладкие мышцы желудка, кишок, матки, мочеточников имеют спонтанную миогенные активность, т.е. развивают спонтанные тетаногиодибни сокращения. Они хранятся в условиях изоляции этих мышц и при фармакологическом выключении интрафузальных нервных сплетений. Итак, ПД возникает в собственно гладких мышцах, а не обусловлен передачей в мышцы нервных импульсов.
Эта спонтанная активность имеет миогенные происхождения и возникает в мышечных клетках, которые выполняют функцию водителя ритма. В этих клетках местный потенциал достигает критического уровня и переходит в ПД. Но за реполяризацию мембраны спонтанно возникает новый местный потонциал, который вызывает еще один ПД, и т.д. ПД, распространяясь через нексус на соседние мышечные клетки со скоростью 0,05-0,1 м / с, охватывает весь мышцу, вызывая его сокращение. Например, перистальтические сокращения желудка возникают с частотой 3 раза за 1 мин, сегментарные и Маятникообразные движения толстой кишки-в 20 раз за 1 мин в верхних отделах и 5-10 за 1 мин - в нижних. Таким образом, гладкие мышечные волокна названных внутренних органов обладают автоматизмом, который проявляется их способностью ритмически сокращаться при отсутствии внешних раздражителей.
Какова причина возникновения потенциала в клетках гладких мышц водителя ритма? Очевидно, он возникает вследствие уменьшения калиевой и увеличение натриевой и (или) кальциевой проницаемости мембраны. Что касается регулярного возникновения медленных волн деполяризации, наиболее выраженных в мышцах ЖКТ, го нет достоверных данных об их ионное происхождения. Возможно, определенную роль играет уменьшение первоначального инактивирующего компонента калиевого тока при деполяризации мышечных клеток вследствие инактивации соответствующих ионных калиевых каналов. Благодаря этому становится возможным возникновение повторных Г1Д.
Эластичность и растяжимость гладких мышц.
В отличие от скелетных мышц, гладкие при растяжении себя как пластичные, эластичные структуры. Благодаря пластичности гладкая мышца может быть полностью расслаблен как в сокращенном, так и в растянутыми состоянии. Например, пластичность гладких мышц стенки желудка или мочевого пузыря по мере наполнения этих органов предотвращает повышение внутриполостного давления. Чрезмерное растяжение часто приводит к стимулированию сокращения, которое обусловлено деполяризацией клеток водителя ритма, возникающий при растяжении мышцы, и сопровождается повышением частоты ПД, а вследствие этого - усилением сокращения. Сокращение, которое активизирует процесс растяжения, играет большую роль в саморегулировании базального тонуса кровеносных сосудов.
Механизм сокращения гладких мышц. Обязательным условием возникновения сокращение гладких мышц, как и скелетных, е увеличение концентрации Са2 + в миоплазми (до 10в-5 М). Считается, что процесс сокращения активизируется преимущественно внеклеточным Са2 +, поступающего в мышечные клетки через потенциалзависимые Са2 +-каналы.
Особенность нервно-мышечной передачи в гладких мышцах заключается в том, что иннервация осуществляется вегетативной нервной системой и она может оказывать как возбуждающий, так и тормозящее влияние. По типу различают холинергические (медиатор ацетилхолин) и адренергические (медиатор норадреналин) медиаторы. Первые обычно содержатся в мышцах пищеварительной системы, вторые - в мышцах кровеносных сосудов.
Один и тот же медиатор в одних синапсах может быть возбуждающих, а в других - тормозным (в зависимости от свойств циторецепторив). Адренорецепторы делят на а-и В-. Норадреналин, воздействуя на а-адренорецепторы, суживает кровеносные сосуды и тормозит моторику пищеварительного тракта, а воздействуя на В-адренорецепторы, стимулирует деятельность сердца и расширяет кровеносные сосуды некоторых органов, расслабляет мышцы бронхов. Описаны нервно-мышечно-. ную передачу в гладких мышцах за помощью и других медиаторов.
В ответ на действие возбуждающего медиатора происходит деполяризация клеток гладких мышц, которая проявляется в виде возбуждающего синаптической потенциала (ССП). Когда он достигает критического уровня, возникает ПД. Это происходит тогда, когда до нервного окончания друг за другом подходят несколько импульсов. Возникновение ЗСГИ является следствием увеличения проницаемости постсинаптической мембраны для Na +, Са2 + и СИ ".
Тормозной медиатор вызывает гиперполяризацию постсинаптической мембраны, что проявляется в тормозном синаптического потенциале (ГСП). В основе гиперполяризации лежит повышение проницаемости мембраны в основном для К +. Роль тормозного медиатора в гладких мышцах, возбуждаемые ацетилхолином (например, мышцы кишки, бронхов), играет норадреналин, а в гладких мышцах, для которых возбуждающих медиатором является норадреналин (например, мышцы мочевого пузыря), - ацетилхолин.
Клинико-физиологический аспект.
При некоторых заболеваниях, когда нарушается иннервация скелетных мышц, их пассивное растяжение или смещение сопровождается рефлекторным повышением их тонуса, т.е. устойчивости к растяжению (спастичность или ригидность).
При нарушении кровообращения, а также под действием некоторых продуктов метаболизма (молочной и фосфорной кислот), ядовитых веществ, алкоголя, усталости, снижения температуры мышц (например, при длительном плавании в холодной воде) после длительного активного сокращения мышцы может возникать контрактура. Чем больше нарушается функция мышцы, тем сильнее выражена контрактурно последействие (например, контрактура жевательных мышц при патологии челюстно-лицевой области). Каково происхождение контрактуры? Считается, что контрактура возникла вследствие уменьшения в мышце концентрации АТФ, что привело к образованию постоянной связи между поперечными мостиками и актиновыми протофибрилл. При этом мышца теряет гибкость и становится твердым. Контрактура проходит, мышца расслабляется, когда концентрация АТФ достигает нормального уровня.
При заболеваниях типа миотонии клеточные мембраны мышц возбуждаются так легко, что даже незначительное раздражение (например, введение игольчатого электрода при электромиографии) обусловливает разряд мышечных импульсов. Спонтанные ПД (потенциалы фибрилляции) регистрируются также на первой стадии после денервации мышцы (пока бездействие не приведет к его атрофии).
Тонические сокращения некоторых гладких мышц, особенно мышц сосудистых стенок (базальный или миогенный, тонус) активизируются преимущественно внеклеточным Са 2 +. Физиологически активные вещества и медиаторы могут вызвать снижение тонуса гладких мышц путем закрытия хемочутливих Са2 +-каналов (через активизацию хеморецепторов) или гиперполяризации, которая обусловливает подавление спонтанных ПД и закрытия потенциалзависимых Са2 +-каналов.
К возбудимым тканям в организме человека относятся нервная, секреторная и мышечная ткани. Однако последняя отличается от остальных уникальным свойством сократимости благодаря наличию в клеточной структуре микрофиламентов из специализированных белков - миозина, актина, тропомиозина.
За счет этого обеспечиваются поддержание позы человека, перемещение в пространстве, продвижение пищевого комка по желудочно-кишечному тракту, кровообращение и многое другое. В зависимости от гистологических особенностей, выполняемых функций и происхождения существует классификация на гладкие и поперечно-полосатые мышечные ткани, также за особенности некоторые авторы выделяют в третий подвид сердечную. Однако следует понимать, сократительные элементы составляют только основу этих тканей, и они не могли бы полноценно работать без густой сети кровеносных сосудов, обеспечивающих доставку большого количества кислорода, защитной и энергетически значимой оболочки - сарколеммы, а также подкрепляющей рыхлой неоформленной
Поперечно-полосатая

Поперечно-полосатые мышечные ткани, в основном, составляют всю скелетную мускулатуру и обеспечивают движение суставов и поддержание позы. Структурно-функциональной единицей их является саркомер, а состоят они, в свою очередь, из миосимпластов - волокон, образовавшихся при слиянии нескольких отдельных клеток в процессе дифференцировки. На гистологическом препарате поперечно-полосатые мышечные ткани легко отличить по многоядерности и исчерченности, отчего они и получили свое название. Другой важной функцией их является теплообразование, в связи с чем при понижении температуры у человека возникает дрожь. Также поперечно-полосатая образует и структуру миокарда, только кардиомиоциты отличаются отсутствием симпластов. При микроскопировании они представляют собой одноядерные веретеновидные клетки. Подразделяются они согласно своим функциям на рабочие (преобладают по количеству), проводящие и секреторные. Именно благодаря вторым, сердечные поперечно-полосатые мышечные ткани обладают свойством автоматизма, то есть имеют способность самостоятельно сокращаться, что и обеспечивает непрерывную работу сердца. Третий же тип клеток является местом синтеза гормоноподобных веществ, в частности это предсердный натрийуретический фактор, способствующий усилению диуреза.
Гладкая
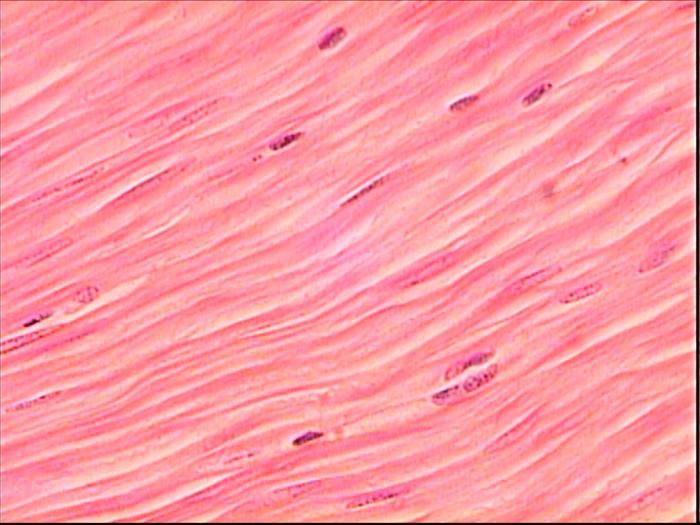
Если обеспечивающая передвижение мышечная ткань человека - поперечно-полосатая, то перистальтику желудочно-кишечного и мочеполового тракта, сокращение сосудистой и бронхиальной стенки обеспечивает гладкая мускулатура. Она отличается ритмичностью, относительной медлительностью, высокой степени растяжимостью и регенераторными способностями, а также вегетативной иннервацией. Это вытянутой формы одноядерные клетки, без исчерченности, и с большим количеством актина и коллагена в структуре. Каждый такой миоцит покрыт тонкой базальной мембраной, а группы - эндомизием из рыхлой неоформленной соединительной ткани.








- Сырники из творога с яблоками
- Как правильно хранить хлеб: несколько полезных советов
- Сталинская высотка на площади восстания
- Жизнь после смерти в христианстве
- Нужно ли поститься перед исповедью, можно ли есть перед исповедью?
- Пигмалион читать онлайн краткое содержание
- Кант, Иммануил – краткая биография
- История представлений о сознании и основные концепции его понимания Платон сознание
- Что такое подлежащее и сказуемое?
- «Измени мышление — и ты изменишь свою жизнь» Б
- Заговоры и ритуалы на деньги в домашних условиях
- Штрафы гибдд: бухгалтерский и налоговый учет Бухгалтерия как закрывать в 1с штрафы гибдд
- Как консервировать кукурузу зернами и початками в банках в домашних условиях на зиму
- Скумбрия на сковороде, тушеная с морковью и луком Как потушить скумбрию с луком и морковью
- Звезда Руси: значение символа
- Значение оберега звезда руси
- Лазарева суббота - приметы и обычаи, что можно и что нельзя делать в Лазареву субботу?
- К чему снится карп. Что значит карп во сне. Приснился карп по Нострадамусу
- Сергей Мамедов, Член Совета Федерации Федерального Собрания РФ от Самарской области: биография, личная жизнь
- Как надо учить грамматику?